第六部:生化学の基礎 酵素,ビタミン,ホルモン
☆ “ホーム” ⇒ “生活の中の科学“ ⇒ “基礎化学(目次)“ ⇒
ここでは,生体物質の代表的な成分の一である酵素に関連し, 【酵素とは】, 【酵素の分類と命名】, 【酵素の所在による分類】, 【補酵素とは】, 【キノン補酵素】, 【ビタミン補酵素】 に項目を分けて紹介する。
酵素とは
酵素( enzyme )
生体で起こる化学反応に対して活性化エネルギーを下げて反応速度を高める触媒や特定の反応だけに作用する機能などを有する分子で,多くの酵素は,生体内で作り出されるたんぱく質(酵素たんぱく質)で構成されている。
酵素たんぱく質は,筋肉たんぱく質や膜たんぱく質などの他の構造たんぱく質と異なり,分子の一隅に活性中心(活性部位)を備えている。
酵素は生命活動における化学反応に関与するため,生化学研究における一大分野である。
酵素の特徴
目的の反応だけに作用する性質(反応選択性)と作用する物質(基質)が特定される性質(基質特異性)とに分けられる。
反応特異性( reaction specificity )
特定の化学反応に対してのみ触媒作用を示す特徴を指す。消化酵素などいくつかの例外を除き,多くの酵素に見られる。
基質特異性( substrate specificity )
例えば,たんぱく質を一般的な酸触媒で加水分解した場合には,活性化エネルギーの低い箇所のペプチド結合から作用するため,たんぱく質の種類によらず分解し,種々の長さのペプチドやアミノ酸が生成する。
一方,ペプチド分解酵素(ペプチターゼ)を用いた場合には,特定の場所のペプチドに作用し加水分解することができる。この特性を基質特異性という。
ペプチダーゼには,ペプチド鎖の内部のペプチド結合を切断できるもの(エンドペプチダーゼ),ペプチド鎖の末端( N 末端又は C 末端)からアミノ酸を一つずつ,又は二つずつ遊離することができるもの(エキソペプチダーゼ)などの基質特異性を持つものがある。
このような機能は,たんぱく質で構成される酵素の立体構造に起因している。古くからは,鍵と鍵穴に例えたモデルで説明されるが,たんぱく質のアミノ酸配列(一次配列)が選択性に寄与していると考えられている。
酵素の構成
酵素の多くは,たんぱく質のみで構成されるが,たんぱく質以外の因子(補因子)を含む複合たんぱく質で構成される酵素もある。
酵素を構成するたんぱく質鎖(ペプチド鎖)は複数本,又は複数種類ある場合がある。各ペプチド鎖は,固有の三次構造(立体構造)を採る。
複合たんぱく質の酵素では,補因子以外のたんぱく質部分をアポ酵素,アポ酵素と補因子とが結合した酵素をホロ酵素という。補因子には,無機イオン,金属含有有機化合物,ビタミンなどの補酵素( coenzyme )がある。
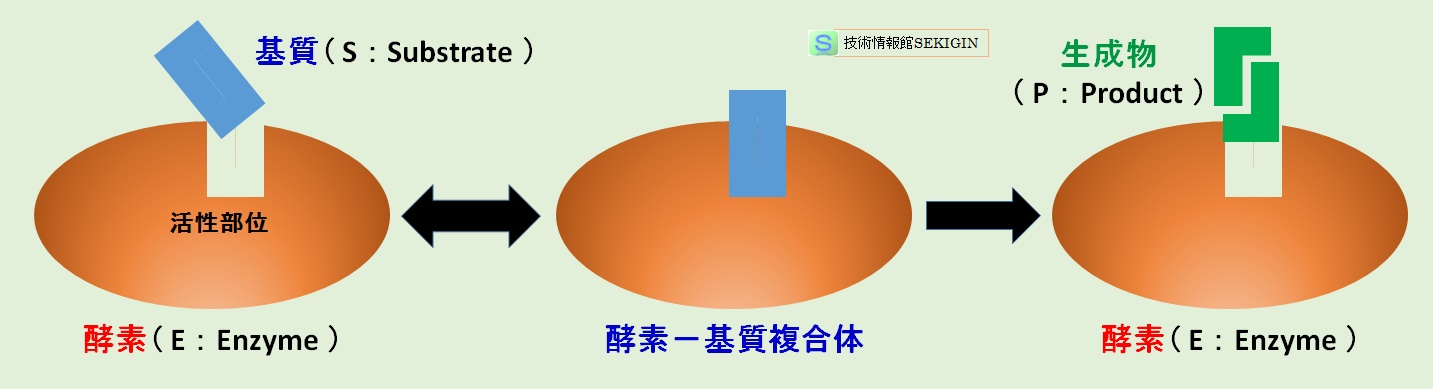
酵素反応(模式図)
ページの先頭へ
酵素の分類と命名
一般的には,酵素の所在による分類が用いられているが,国際生化学分子生物学連合( International Union of Biochemistry and Molecular Biology :IUBMB )では,国際純正・応用化学連合( IUPAC )との協力で,次に示す 6種の 反応形式に従う系統的分類を提唱し,それぞれに EC番号の付与と命名を行っている。
① 酸化還元酵素(オキシドレダクターゼ,oxidoreductase )
EC 第 1 群に分類され,酸化還元反応を触媒する。
② 転移酵素 (トランスフェラーゼ, )
EC 第 2 群に分類され,一方の基質から他方の基質への原子団の移動反応を触媒する。
③ 加水分解酵素 (ヒドロラーゼ,hydrolase )
EC 第 3 群に分類され,加水分解反応を触媒する。
④ リアーゼ ( lyase )
EC 第 4 群に分類され,脱離反応による二重結合生成,逆反応の付加反応で二重結合部位に置換基を導入する反応を触媒する。シンターゼ( synthase ),脱離酵素や除去付加酵素とも呼ばれる。
⑤ 異性化酵素 (イソメラーゼ,isomerase )
EC 第 5 群に分類され,分子内反応を触媒する。
⑥ リカーゼ ( ligase )
EC 第 6 群に分類され,高エネルギー化合物( ATP など)の加水分解と共に触媒作用を示す。シンテターゼ( synthetase ),合成酵素とも呼ばれる。厳密には,合成酵素という場合は EC6 群の他に EC4 群(リアーゼ)を含む。
系統的分類での酵素名には,「系統名」と「常用名」がある。
これらは,IUBMB の定める命名法に従って命名され,EC 番号が与えられる。
系統名は,基質分子の名称(複数の場合は併記),反応の名称を連結して命名される。常用名も系統名と同じ規則で命名されるが,古くに発見され命名された酵素は,当時の名称がそのまま使用されている。
ページの先頭へ
酵素の所在による分類
酵素は,生物体内の代謝反応の関与する場所に存在している。従って,酵素は,その所在により分類することができる。
所在による分類は,大きく,細胞膜や細胞小器官の膜(生体膜)に結合する膜酵素,生体膜以外の細胞質や細胞外に分散している可溶型酵素に分類される。
膜酵素( membrane enzyme )
生体膜に結合している酵素で,膜結合型酵素ともいわれる。
膜酵素は,クエン酸回路で紹介したようなエネルギー授受,物質移動など生体膜の機能を担う酵素で,ATP アーゼ( ATP を ADP とリン酸に加水分解),ATP合成酵素( ADP とリン酸から ATP を合成),チャンネル(無機イオンの輸送体),トランスポーター(たんぱく質の輸送体),ヒドロゲナーゼ(酸化還元反応の触媒)などがある。
これらの膜酵素は,生体膜との結合形態により,酵素の一部が付着している形態の付着型,酵素の一部が生体膜に埋没している埋没型,生体膜を貫通している貫通型に分けられる。
可溶型酵素( membrane enzyme )
たんぱく質である酵素は,他のたんぱく質と同様に,細胞内で合成される。
この中で,水への溶解性が高い物を可溶型酵素といい,細胞質に分散して留まる酵素と合成後に細胞外に分泌されるものに分けられる。
可溶型酵素は,外部には親水性アミノ酸,内部に疎水性アミノ酸が集まる球形構造のものが多い。なお,可溶型酵素の中で,細胞外に分泌される酵素を分泌型酵素と呼ばれる。
細胞質に留まる可溶型酵素は,細胞質での代謝に関わり,分泌型酵素は,消化酵素など,細胞外の物質に作用し,細胞内への取り込みが可能な物質に変える役割のものが多い。
ページの先頭へ
補酵素とは
補酵素( coenzyme )
コエンザイム,助酵素,コエンチームなどともいわれる。酵素の触媒反応で,補助的な役割を果たすある特定の物質の中で,たんぱく質部分(アポ酵素)と可逆的に結合(結合が弱く解離しやすい)する補因子をいう。
補因子( cofactor )
酵素の触媒活性に必要なたんぱく質以外の化学物質(分子,イオン)で,酵素とゆるく結合し,酵素反応の通常の段階では解離される補酵素と酵素のたんぱく質との結合が強く,常時結合し酵素の一部である補欠分子団( Prosthetic group )に分けられる。補欠分子団には,有機物(ビタミン,糖,脂質など)の他に無機物(金属イオンなど)がある。
補酵素は,低分子量の有機化合物で,キノン補酵素,ビタミン補酵素,ATP などで,補因子以外のたんぱく質のアポ酵素と結合した複合体(ホロ酵素)として酵素作用を示すようになる。
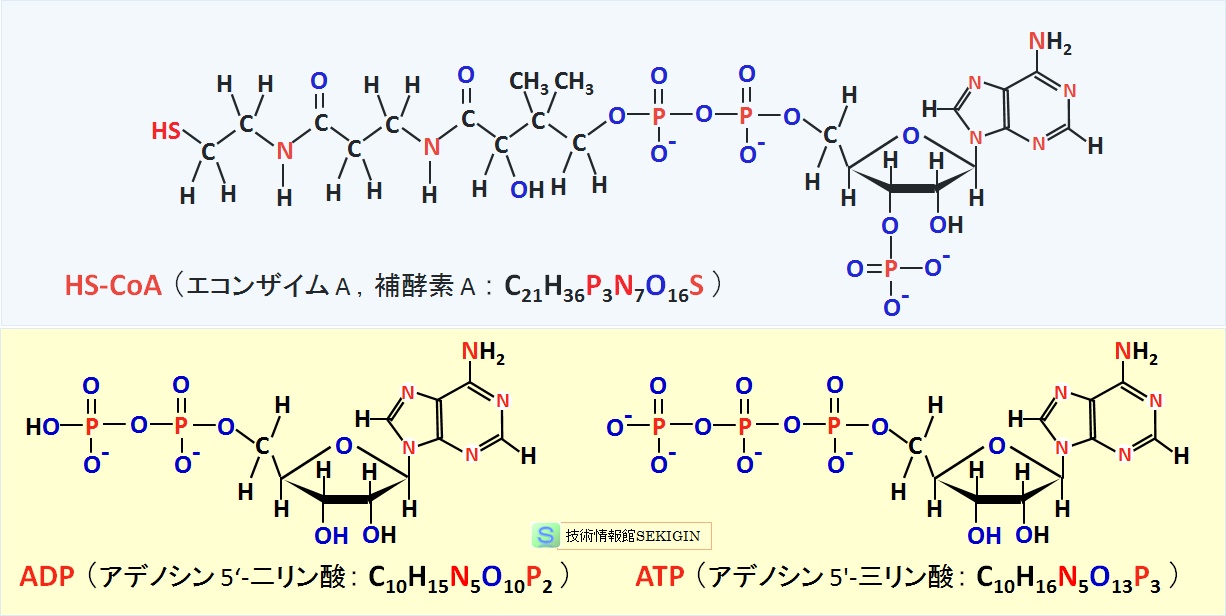
主要な補酵素(補酵素 A ,ADP ,ATP )
ページの先頭へ
キノン補酵素
キノン( quinone )
一般的には,ベンゼン環から誘導されたベンゾキノンなど,2 つのケトン(>C=O )を持つ環状の有機化合物をいう。
主なキノン補酵素を次に紹介する。
PQQ(ピロロキノリンキノン,pyrroloquinoline quinone )
メタノール脱水素酵素やグルコース脱水素酵素に含まれる酸化還元補酵素で,抗酸化作用や神経保護作用などを有する。
TPQ(トパキノン,topaquinone )
アミン酸化酵素(酸化的脱アミノ反応)の補酵素で,前駆体たんぱく質中のチロシン残基が銅イオンの存在下で自動的に酸化されて生成したビルトイン補酵素の一種である。
TTQ(トリプトファン-トリプトフィルキノン,tryptophan tryptophylquinone)
メチルアミン脱水素酵素(メチルアミン酸化還元)の補酵素で,ビルトイン補酵素の一種である。
LTQ(リシンチロシルキノン,lysine tyrosylquinone )
ペプチジルリシン酸化酵素(ペプチド内リシンの酸化)の補酵素で,ビルトイン補酵素の一種である。
CTQ(システニル-トリプトファンキノン,cysteine tryptophylquinone )
キノヘムプロテイン・アミン脱水素酵素(QHNDH)に含まれる補酵素で,ビルトイン補酵素の一種である。
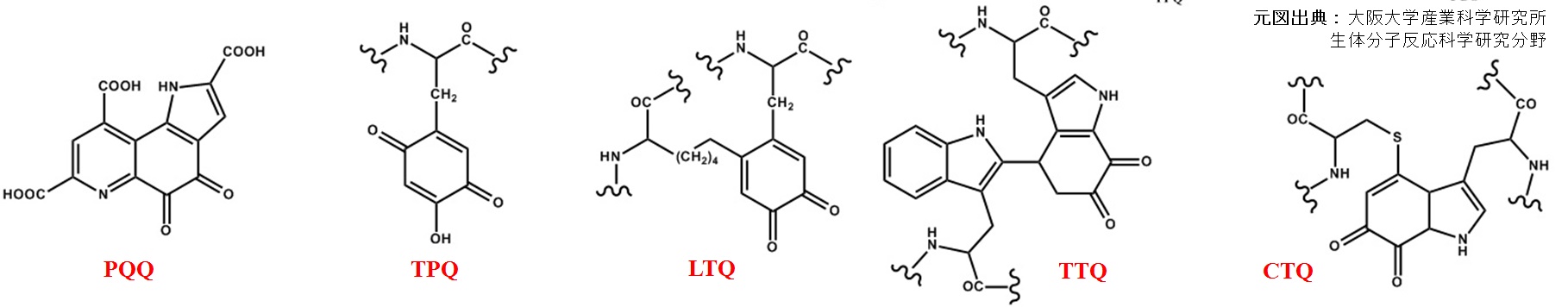
キノン補酵素
元図出典:大阪大学産業科学研究所QHNDH
ページの先頭へ
ビタミン補酵素
水溶性ビタミンから誘導される補酵素で,主なものを次に紹介する。
補酵素 A(コエンザイム A ,coenzyme ,CoA )
糖の代謝や脂肪酸の代謝などの重要な反応に関わる補酵素で,パントテン酸( pantothenic acid )と呼ばれるビタミン B5 ,アデノシン二リン酸,2-チオキシエタンアミンから構成(化学式: C21H36P3N7O16S )される。
アセチル基が結合したアセチル CoA など多くの補酵素 A の誘導体がある。主な誘導体を挙げると,アセトアセチル CoA ,カフェオイル CoA ,グルタリル CoA ,クロトニル CoA ,シナポイル CoA ,シンナモイル CoA ,スクシニル CoA ,3-ヒドロキシブタノイル CoA ,ヒドロキシメチルグルタリル CoA ,フェルロイル CoA ,プロピオニル CoA ,マロニル CoA などがある。
NAD+,及び NADP+
NAD+ ,NADP+ は,ナイアシン( Niacin ),ビタミン PP とも呼ばれるビタミン B3 から誘導され,約 500 種類の酸化還元酵素の補酵素として機能する。
NAD+(ニコチンアミドアデニンジヌクレオチド, nicotinamide adenine dinucleotide )は,全ての真核生物の他に多くの菌類で用いられる電子伝達体で,酸化型の NAD+ と還元型の NADH が,可逆的に相互変換できる。
NADP+(ニコチンアミドアデニンジヌクレオチドリン酸,nicotinamide adenine dinucleotide phosphate )は,光合成経路,解糖系のエントナー-ドウドロフ経路などで用いられる電子伝達体で,酸化型の NADP+ と還元型の NADPH が,可逆的に相互変換できる。
FAD,及び FMN
FAD ,FMN とも,ビタミン B2(リボフラビン)から誘導される。
FAD(フラビンアデニンジヌクレオチド,flavin adenine dinucleotide )は,フラボ酵素やフラビンタンパク質と呼ばれる酸化還元酵素において,電子移動で機能する補酵素である。
FAD →(還元)→ FADH2 , FADH2 →(酸化)→ FAD
FADH2 は,ミトコンドリアでの酸化的リン酸化の基質として使われる。FADH2 は,酸化されて FAD になるとき,ATP を 2 分子作ることができる。
FMN(フラビンモノヌクレオチド,flavin mononucleotide )は,多くの酸化還元酵素の補酵素である。FMN は,FAD と同様に,酸化型の FMN と還元型の FMNH2 が,可逆的に相互変換できる。
補酵素 F( THF )
葉酸( folate ),ビタミン M ,プテロイルグルタミン酸などと呼ばれるビタミン B9 は,小腸細胞のジヒドロ葉酸レダクターゼによってプテリン環が還元されて補酵素のテトラヒドロ葉酸( Tetrahydrofolic acid : THF )に変換される。種々の C1 基の転移反応,特にアミノ酸の代謝と核酸の代謝に関わる補酵素として作用する。
TPP
TPP(チアミンピロリン酸,thiamine pyrophosphate )は,チアミン二リン酸( thiamine diphosphate :ThDP )ともいい,ビタミン B1 のリン酸化で得られ,トランスケトラーゼ,ピルビン酸脱水素酵素,α-ケトグルタル酸脱水素酵素などの補酵素である。
PLP
PLP(ピリドキサールリン酸,pyridoxal phosphate )は,ピリドキサール( pyridoxal ),ピリドキシン( pyridoxine ),ピリドキサミン( pyridoxamine )などと呼ばれるビタミン B6 から誘導される補欠分子族の一つである。
PLP は,栄養素の代謝,神経伝達物質合成,ヒスタミン合成,ヘモグロビン合成などの多くの反応に関与する酵素(アミノ基転移,CO2 離脱)と共に機能する。
補酵素 R(ビオチン)
ビオチン( biotin ),ビタミン Bw ,ビタミン H などと呼ばれるビタミン B7 は,カルボキシル基転移酵素( carboxylase )の補酵素として二酸化炭素の転移を触媒する。特にビオチンを補酵素として持つ酵素の一群をビオチン酵素( biotin enzyme )と呼ぶ。
補酵素B12(ビタミンB12)
シアノコバラミン( cyanocobalamin ),ヒドロキソコバラミン( Hydroxocobalamin )などと呼ばれるビタミン B12 は,細胞に入り補酵素として働く活性型(メチルコバラミン,アデノシルコバラミン)に変換し,水素移動,メチオニンの生成でアミノ酸や脂肪酸の代謝および葉酸の生合成に用いられる。
ページの先頭へ